
董梦秋:化学交联质谱让结构生物学研究如虎添翼
发布时间:2018-10-31 阅读次数:10565次
在蛋白质组学分析方法中,质谱获得的是多肽序列结构的信息;那么用质谱是否可研究大分子蛋白的结构信息?近几年来,董梦秋实验室在中国做出了多项先驱性工作,主要集中在化学交联质谱领域。在用单颗粒冷冻电镜技术研究结构生物学屡创佳绩的当下,很多研究者都把样品一分为二,一份做冷冻电镜,一份做交联质谱。那么交联质谱如何让结构生物学研究如虎添翼?在中国质谱学术大会召开前夕,应学会秘书处邀请,分析测试百科网采访到北京生命科学研究所的董梦秋研究员。她将为我们介绍交联质谱新方法,也对中国质谱大会的召开表达了衷心的祝愿。

北京生命科学研究所 董梦秋研究员
质谱大会开启学术交流瘦身健体的新时代
谈到2018年中国质谱学术大会的意义,董梦秋认为其意义重大。她说:“物理学会和化学学会的质谱会议合二为一,诞生了2018年中国质谱学术大会,开启了学术交流瘦身健体的新时代。当前科研人员会议负担过重,以至于有人感叹‘不是在开会,就是在去开会的路上,’没有足够的时间沉静下来做科研。学术会议太多,表面上看热热闹闹一片繁荣,实则干扰到正常工作。中国科研就像一个青春期的孩子,快速成长,但也有虚胖的成分。瘦身健体,减负前行,更有利于发展。”
pLink和交联质谱在国际上的影响力
与化学交联质谱的结缘
董梦秋曾在著名蛋白质组学大师John Yates教授实验室做博士后,Yates教授最享誉全球的工作是开发了SEQUEST蛋白质组学“鸟枪法”鉴定软件。当时,有位合作者研究一种HSP90的复合体,认为其结合方式同以前报道的不同,希望能用化学交联质谱(CXMS)判断复合体中两个蛋白的结合界面。那时只有常规肽段鉴定工具,董梦秋用18O标记区分交联肽段和普通肽段,通过大量搜索和辅助人工判断,终于完成了任务。虽然研究很有意思,但因为没有专用软件工具,做起来很费劲。
董梦秋2007年到北京生命研究所(NIBS)做PI,找到了中科院计算所贺思敏教授,从此董梦秋开始了与pFind 团队的长期合作。他们致力于开发交联质谱技术,打造出已经成为业界标杆的pLink软件和一系列工作流程。
交联质谱的价值
董梦秋谈到,“近年来质谱技术在结构生物学领域的重要性愈发彰显。现在结构生物学家研究的蛋白质复合体越来越大,种类、状态越来越多,即使有强大的电镜和晶体学手段也不一定能看清所有的关键性的结构细节、状态间的差异,或者不能确定组成亚基的化学计量比,他们的需求带动了交联质谱、native MS和氢氘交换等技术的应用和发展。看看结构文章的作者名单就知道,很多都有质谱专家的身影。”
董梦秋实验室和pFind 团队的合作开展不到一年,Ruedi Abersold团队就在Nature Methods上发表了他们开发的交联质谱鉴定软件,但该软件有一个明显的缺陷:没有假阳性率的估算,无法判断鉴定结果的好坏。2012年董梦秋实验室和pFind团队合作开发的pLink软件在《Nature Methods》发表。在这项工作中快速寻找候选肽、谱图打分、评价、控制假阳性率等环节一步到位。该软件和配套的方法流程还有一大好处:不要求必须使用稳定同位素标记的交联试剂,仅需使用便宜上千倍的普通交联试剂。迄今,pLink在全球有800多个用户,被众多新开发者当作CXMS软件的业界标杆来比较。
交联质谱让结构生物学研究如虎添翼
质谱研究结构生物学的方法
在介绍交联质谱前,董梦秋为我们普及了用质谱研究结构生物学的方法。在当今的结构生物学研究中,冷冻电镜是主力。从单颗粒冷冻电镜图像中把形态相近的颗粒挑出来进行对齐、平均、三维重构,可以得到高分辨度的电子密度图,然后搭建蛋白质复合体的结构模型。可是,蛋白质柔性区域的结构无论用电镜还是晶体学技术都很难看清楚,甚至完全不可见,这样建模的时候会遇到断头路,往下接到哪儿是个问题。交联质谱数据告诉我们,两个被交联的氨基酸残基空间距离不会超过某个上限。有了足够多的交联距离约束信息后,我们就可以把断头路接起来,并推断出柔性区的位置和分布方式。目前很多结构生物学实验室在制备冷冻电镜样品时会分出一部分做交联质谱,后期匹配两种数据以获得更完整的结构。而Native MS(非变性质谱)可以提供蛋白质复合体组成亚基的化学计量比,帮助搭建合理的结构模型。Foot Printing(足迹法)用化学试剂标记暴露在环境中的氨基酸残基,酶解后可用质谱检测被修饰的肽段,从而了解哪些氨基酸分布在蛋白质的外表面以及蛋白质形成复合体后被包埋的区域。交联质谱数据中的mono-link肽段(交联剂一端与肽段共价连接,另一端水解而形成的修饰单肽)也可以提供同类信息。氢氘交换不受化学修饰试剂的选择性限制,可以做得更精细。
交联质谱的基本原理
研究蛋白质结构与相互作用的传统技术,如核磁共振技术、X 射线晶体衍射技术等,对于蛋白质的纯度、结晶性和绝对量均有比较高的要求,限制了其广泛应用。化学交联结合质谱技术(chemical cross-linking of proteins coupled with mass spectrometry),简称交联质谱技术、CXMS或XL-MS,是近年发展起来的新方法。它利用化学交联剂 (chemical cross-linker) 处理蛋白质样品,将空间距离足够接近、可以与交联剂反应的两个氨基酸以共价键连接起来,然后利用基于高精度质谱的蛋白质组学手段分析交联产物。交联反应发生在溶液态,反映了蛋白的溶液构象。
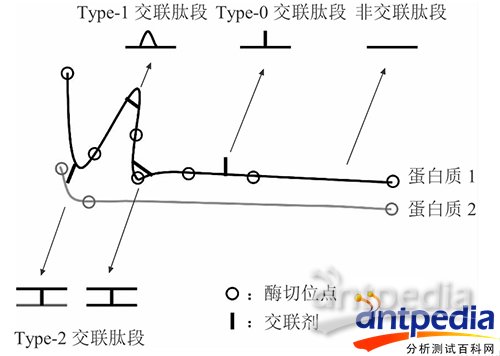
交联蛋白质酶切后产生的交联肽段的各种形式
交联蛋白质酶切后可以产生三种交联产物(如上图):a.被交联剂修饰的单肽 (交联剂只一端交联一条肽段,mono-linked peptide),Type-0 类型的交联肽段;b.肽段内部交联 (交联剂两端交联同一条肽段,loop-linked peptide 或intra-linked peptide),Type-1类型的交联肽段;c.肽段间交联 (交联剂交联两条肽段,Inter-linked peptides),Type-2 类型的交联肽段。其中,最有用的是Type-2类型。
交联质谱的应用实例
第一个例子,同UCLA一个实验室合作研究mTORC1激酶复合体以何种方式募集底物蛋白4E-BP1(J Biol Chem. 2014 Feb 21.)。mTORC1激酶复合体由mTOR、Raptor和LST8三个蛋白组成。有人认为募集4E-BP1的是mTOR,也有人认为是Raptor。mTORC1与4E-BP1的交联质谱结果表明只有Raptor N-端的第一个RNC结构域同4E-BP1间有交联。
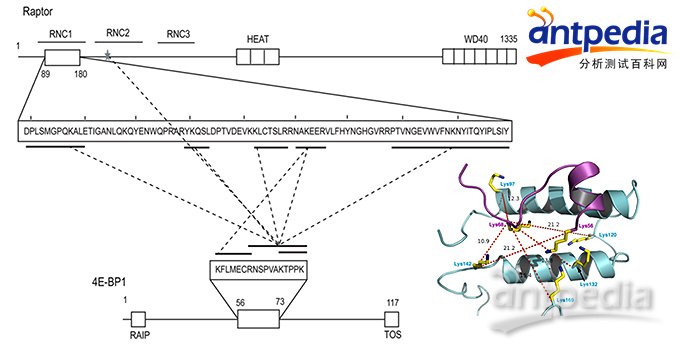
用交联质谱揭示Raptor和4E-BP1的作用界面信息
依据交联结果的距离约束信息构建结构模型,再用肽段竞争实验进行生物学验证,可以确认交联质谱结果可靠,mTORC1复合体中的Raptor亚基招募了4E-BP1。 如上图所示,Raptor两个α-螺旋(α-helix) 形成的“基座”与4E-BP1肽段(紫色)结合
第二个例子,同NIBS一个遗传学实验室合作,研究裂殖酵母中两个水解酶APE2和LAP2是如何通过一个叫NBR1的蛋白运送到溶酶体中的(Mol Cell. 2015 Sep 17.)。利用交联质谱发现APE2是关键,因为另外两个蛋白都与它的N末端有交联。去掉APE2 N端的5个氨基酸后再做实验并用荧光标记,证实了该结果。没有这些N端的关键氨基酸结合位点,就不能形成三元复合体,两个水解酶都无法进入溶酶体中。
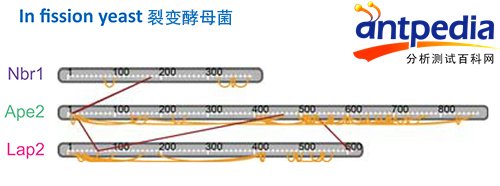
Nbr1、Ape2和Lap2的结合
在第三个例子中,交联质谱与冷冻电镜技术结合,解析核糖体前体的组装过程(Nature, 2016,细胞核内的核糖体组装前体结构揭示了装配成熟因子的功能多样性),合作者是清华大学高宁教授。核糖体前体在加工过程中,有很多组装因子(Assembly factor)蛋白参与,组装任务完成后它们被去除。比如,有很多组装因子结合在pre-60S核糖体前体的ITS2区域,但它们的结构未知(不知道积木块儿长什么样),不能用搭积木的方法归属该区域的电子密度。通过二级结构搜索,比如把电镜看到的长长短短的α-螺旋与二级结构预测的α-螺旋进行比对,找到部分肽段的位置,再加上交联质谱信息一点点推断、确认,最后全部解析这部分结构。
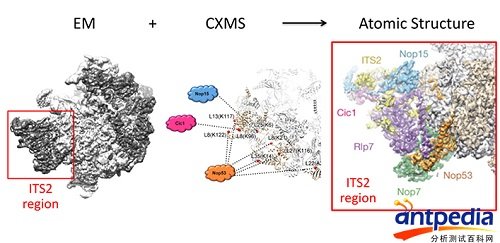
用冷冻电镜+交联质谱揭示核糖体组装前体结构
第四个例子是同清华大学王宏伟教授的合作研究(J Mol Biol., 2017)。WHAMM蛋白的作用是沿着微管铺就的轨道运送囊泡(vesicle)。它一端结合囊泡,另一端结合微管。与微管结合后,WHAMM的微管结合域 (MTBD) 在冷冻电镜中只能看到被称为MBM(Microtube Binding Motif,图中用紫色表示)的一小部分,它与微管紧密结合;其余部分(图中用黄色表示)因为高度灵活(flexible),所以电镜无法看清。通过交联质谱发现, MBM(紫色部分)在没有结合微管时与MTBD的其它区域(黄色)有大量交联,但结合了微管后就看不见了。该结果与冷冻电镜结果互补,启发我们得到一个合理的解释,即MTBD结合微管前后发生了巨大的构象变化,MBM与MTBD的其余部分原本紧挨着,一旦结合了微管两边就分开了,而且相互之间可以随意摆动。
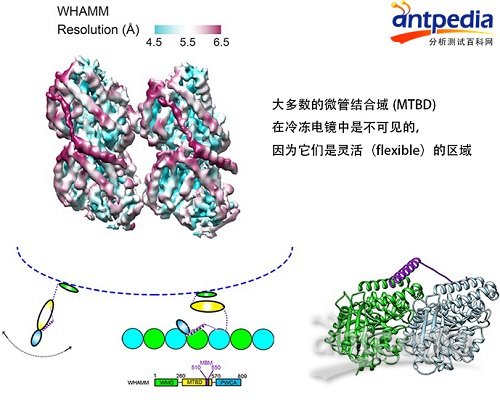
用交联质谱揭示WHAMM和微管结合的、冷冻电镜看不清的Flexible区域
交联质谱还可以鉴定体内天然发生的一些交联,如二硫键。关注蛋白质药物研发的生物制药公司有很多这方面的需求。交联质谱还可以表征蛋白质动态。董梦秋实验室与武汉物理与数学研究所唐淳实验室合作在JBC发表题为“Modeling protein excited-state structures from "over-length" chemical cross-links”的研究论文,成功开发了基于交联质谱的研究蛋白质动态 (protein dynamics) 的新方法,并发布了通用流程。通过交联数据和结构计算发现,结合了钙离子但没有结合配体肽段或蛋白的钙调蛋白(Ca2+-CaM) 除了采取哑铃状的开放态构象(基态),还有一种闭合态构象,非常接近之前用NMR手段观察到的激发态构象。文章的艺术配图被选为当期JBC封面(J Biol Chem. 292(4):1187-1196. 2017)。

尽量减少会议 积极分享
谈到自己如何支持中国质谱的发展,董梦秋谈到几点:首先,支持学术会议做减法,合并同类项;其次是分享,把实验室的技术分享给其他人。作为今年8月第五届中国计算蛋白质组学研讨会(CNCP-2018)的一部分,董梦秋实验室同pFind 团队一起举办了第二届交联质谱分析实验培训,用三天的时间手把手培训了来自来全国各地的十位学员。他们自己动手制样、上机、分析数据,完整体验每一个环节。“我们把经验都分享给他们,效果很好。大连化物所许国旺老师实验室10月份举办代谢组学高级培训班,给大家提供了很好的学习机会,我们也有学员报名参加。类似这样的开放分享我觉得很好。”
让质谱服务于各学科各领域 质谱将更加有影响力
关于中国质谱在世界上的影响力和地位,董梦秋说,“关注这个问题有点儿虚”,质谱技术是个工具,它要服务的对象是生物、化学、物理、医学等学科。把这些“用户学科”的需求放在首位,关注如何去满足它们的需求,尽力帮助它们解决问题,质谱技术自然就获得了推动力,也必然带动化学、物理、工程和计算科学等相关领域的研究,不断推进质谱技术本身的发展。进入这个正循环,中国质谱在世界上的影响力和地位自然就越高。总之,努力使自己更加有用,服务于他人;越有用,越重要,就越有影响。
最后,董梦秋预祝2018年中国质谱学术大会圆满成功! 她由衷表达:为中国质谱新时代、新风尚点赞!向幕后的促成者、组织者致敬!
人物简介:
董梦秋博士,北京生命科学研究所研究员。董梦秋研究员早年毕业于四川大学,1995年硕士毕业于中科院上海生化所,2001年博士毕业于耶鲁大学,曾经在美国加州大学圣地亚哥分校、斯克利普斯(Scripps)研究院从事博士后研究,2007年至今,北京生命科学研究所研究员。董梦秋实验室的研究主题包含生物学和质谱技术两部分,二者相辅相成,共同发展。在生物学研究方面,以线虫为模式生物研究衰老过程及其调控机制;对线虫自然衰老过程中亚细胞器(如线粒体)和分子水平(转录组和蛋白质组)的变化进行分析,比较长寿突变体线虫与野生型线虫之间有何不同。在衰老的调节机制的研究中,主要关注胰岛素信号通路。在质谱方面,致力于基于质谱的蛋白质组学技术的应用与开发,主要包括肽序列从头测序的算法开发、基于电子转移解离(ETD)谱图的高效蛋白质鉴定技术、和通过鉴定交联肽段来研究蛋白-蛋白相互作用的方法。实验室目前有四台质谱仪,包括一台装备了ETD的LTQ-Orbitrap,两台 Q Exactive,以及一台 Orbitrap Fusion LUMOS。